Популяционное деление европейского лося
ПОПУЛЯЦИОННОЕ ДЕЛЕНИЕ ЕВРОПЕЙСКОГО ЛОСЯ (AlcesalcesalcesL.) И ПРОБЛЕМА АНТРОПОГЕННОГО РАЗДРОБЛЕНИЯ ЕГО АРЕАЛА
А.В. Давыдов1, М.В. Холодова2,И.Г. Мещерский2, С.А. Царев3, Ю.П. Губарь1, А.Б. Линьков3, А.В. Проняев1, Н.Э. Овсюкова1, Л.В. Рожкова1, Ю.И. Рожков1.
1ФГУ “Центрохотконтроль” МСХ РФ, 2Институт проблем экологии и эволюции им. А.Н. Северцова РАН, 3 ФГОУ ВПО«Российский государственный аграрный заочный университет»
Популяцию кратко можно определить, как группировку особей одного вида в той или иной степени изолированной от других подобных группировок. Мы выделяем следующие иерархически организованные группировки:
- мегапопуляция — структура, охватывающая в одних случаях вид в целом, в других - наиболее крупные внутривидовые группировки (уровень полувида-подвида);
- макропопуляция — структура, включающая единицу меньшей масштабности, соответствующей чаще всего географической или экологической расе;
- мезопопуляция — группировка, входящая в состав макропопуляции, более или менее ограниченная пространственно и частично изолированная “преградами” или же расстоянием от других подобных группировок;
- микропопуляция ― группировка, ограниченная территориально и далее не делимая на популяционные единицы внутри мезопопуляции.
Когда необходимо увеличить детализацию, мы вводим “цифровую номенклатуру” в тот уровень, который дополнительно нами подразделяется (например, мегапопуляция Iи IIрангов) макропопуляция IIIи IVпопуляционных рангов.
Население лося Европы можно представить себе как единую макропопуляцию высшего ранга, простирающуюся на севере от побережья Баренцева и Белого морей и доходящую на юге почти до северного Кавказа. На западе она ограничена современным ареалом лося на этих территориях, т.е. лежит относительно близко к границам бывшего Советского Союза и, пожалуй, еще ближе к границам Российской империи, когда в ее состав входили Финляндия и Польша. На востоке эта европейская макропопуляция, в районе Уральского хребта, плавно переходит в другую макропопуляцию – Западно-Сибирскую, которую здесь мы рассматривать не будем.
В свою очередь, европейскую макропопуляцию можно разбить на две дополнительные макропопуляционные группировки более низкого ранга, чем предыдущая - североевропейскую и южно-европейскую.
Типичным местообитанием первой являются северо- и среднетаежные леса, второй — лесостепные и степные территории (вернее отдельные лесные массивы на этих территориях).
Обе эти макропопуляции соединяются в центральной полосе России переходной зоной шириной около 300 км. Эта зона охватывает южнотаежные, смешанные и широколиственные леса (рис.1).
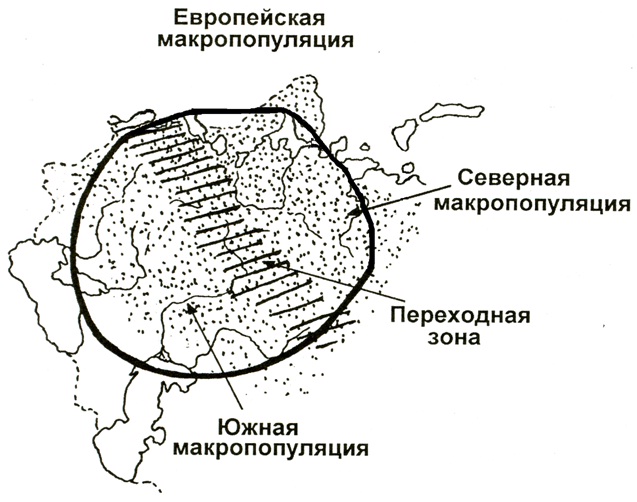
Рис.1. Ареал лося (показан точками) на территории РФ с наложением на него схематичным обозначением макропопуляционных группировок и переходной зоны (штриховка)
Макропопуляции в свою очередь, как мы уже отмечали, можно разделить на группировки еще меньшего масштаба — мезопопуляции. Их "границы" друг от друга, конечно же, условны, как и предыдущих, более крупномасштабных, группировок. Однако, если в первом случае, скорее всего, существенную роль играет изоляция расстоянием и "пристрастие" особей к определенным биотопам, то во втором, по-видимому, важнее притормаживающие потоки генов преграды (например, естественные — обширные водоемы, крупнейшие реки и искусственные - территории, населенные человеком или, в тех или иных формах, эксплуатируемые им своего рода "репеллентные" ландшафты).
Вполне понятно, что не только северная и южная макропопуляции, но и вся переходная зона заполнена мезопопуляциями.
Каждая из подобных группировок, хотя и не может быть четко ограничена, скорее всего, занимает площади от 25 до 100 тыс. кв. км, т.е. пространство, соответствующее по площади одной - четырем областям центрального района (это как раз те площади, которые могут "уложиться" между наиболее значимыми препятствиями).
Мезопопуляции, делимые на микропопуляционные группировки, занимают территории, лежащие в пределах 100 — 200 кв. км. (естественно, с некоторыми вариациями пределов).
Количество особей, держащихся на такой территории и составляющих собственно микропопуляцию, колеблется от 25 до 250 особей (опять-таки с возможными вариациями пределов)
Микропопуляции разбросаны по макропопуляционному пространству неравномерно. В переходной зоне, на 30 тыс. кв. км. (эта цифра приблизительно соответствует среднему размеру области Центрального региона РФ), приходится порядка 100 микропопуляций. На севере, на ту же территорию, их приходится гораздо меньше - около 20. И еще меньше на юге - около 10 микропопуляций.
Расстояния между популяционными ядрами следующие. Переходная зона 20 км (плюс- минус 10 км), Север -50 -100 км, Юг - 50-200 км (что соответствует расстояниям между лесными массивами в степи — лесостепи).
Таким образом, "минимальные" популяции распределены мозаично - обитаемые территории чередуются с незаселенными пространствами, которые наиболее обширны на севере и юге. Последние не заселены не только потому, что они "цивилизованы" человеком или экологически не пригодны для вида, но и в силу других причин.
Теперь коснемся морфологических различий между популяциями (точнее, различий в массах тела). Начнем опять с макропопуляций.
Если сравнить северных особей с соответствующими по полу и возрасту южными, то выясниться, что они в среднем на 10 % тяжелее. Если же посмотреть, как изменяются значения средней массы в европейской макропопуляции по трансекте, проложенной с севера Архангельской области до Белгородской на юге, то выявится интересная закономерность. Массы на юге и севере неизменны, они меняются скачком лишь в переходной зоне (рис.2). Подобным же образом ведут себя и другие морфологические характеристики (табл.1).
Таким образом, северная и южная макропопуляции на всем пространстве занимаемых ими территорий совершенно "самобытны" в морфологическом плане. Они "контактируют" в переходной зоне, являющейся по существу зоной интрогрессии, т.е. той территорией через которую перетекают гены (в виде особей, конечно), из одной популяции в другую, в результате чего и создается 300 километровый градиент масс.
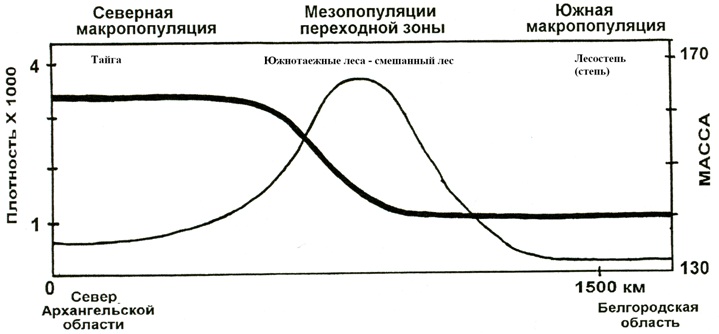
Рис. 2. Изменение массы (широкая линия) лосей с севера на юг и их плотности (тонкая линия). Точки значений масс и плотности на сглаженных кривых опущены, подробно информацию можно найти в других наших публикациях (Рожков и др., 2001; Давыдов и др., 2004).
Таблица 1
Характеристика рогов лосей Европейской территории РФ на основе оценки охотничьих трофеев (Давыдов и др., 2004)
Параметр | Северная макро- популяция n=105 | Переходная зона
n=116 | Южная макро- популяция n=51 |
Окружность, см | 17,98±0,2 | 17,54±0,13 | 16,7±0,22 |
Развал, см | 112,9±1,42 | 108,32±1,13 | 105,61±1,67 |
Длина, см | 97,9±0,97 | 98,41±0,81 | 94,71±1,4 |
Ширина лопаты, см | 24,5±0,54 | 21,95±0,5 | 17,91±0,64 |
Оценка в баллах | 302,0 | 287,5 | 268,4 |
Доля оленеобразных рогов, % | 4,8±2,09 | 9,5±2,72 | 17,6±5,33 |
Масса туши*, кг | 172,8 | 161,7 | 151,4 |
*Примечание: для массы туши значение ошибки не приводится, т.к. выборки содержат тысячи и десятки тысяч особей.
Возникает вопрос: какие силы поддерживают ступенчатый характер этого градиента? При отсутствии этих сил ступень также отсутствовала бы, даже если сделать далекое от реальности предположение, что первоначально особи, заселявшие "юг" с середины XX века были значительно мельче своих северных собратьев. Не было бы ступени, но был бы плавный переход масс и в том случае, если бы действовали естественные силы в виде, например, температуры, которая ответственна за выполнение правила Бергмана (где холодней, там больше масса у теплокровных животных). Как известно, метеопараметры изменяются относительно плавно с севера на юг. Столь же плавно должна была бы меняться и масса.
Единственная сила способная создать ступенчатый градиент имеет искусственный характер - это пресс охоты
Действительно, если наложить друг на друга две карты, на одной из которых отмечена плотность людского населения, а на другой распределение средней массы по территории РФ, то окажется, что высокие плотности населения почти точно совпадут с низкими массами. Причем, изменение плотности с севера на юг имеет точно такой же ступенчатый характер, как и массы. С той лишь разницей, что в 300 километровой переходной зоне совершается зеркально отображенный скачек (так как численность населения мала на севере и велика на юге), который без труда можно себе представить, взглянув на рисунок 2 и дорисовав в воображении симметричную к кривой массы кривую плотности.
Вполне понятно, что плотность охотников пропорциональна плотности населения.
Чтобы окончательно понять механизм образования "ступени" необходимо принять, что изъятие животных по массе не случайно, а выборочно, т.е. в первую очередь изымаются наиболее крупные особи
Теоретические расчеты показывают, что за более, чем полувековой период существования южной макропопуляции (ранее на этой территории лоси практически не встречались) возможен даже больший, чем 10%, сдвиг в массе. Детали этого механизма подробно рассматриваются в нашей книге (Рожков и др., 2001). Там же показано, что даже при отсутствии браконьерства и изъятии животных согласно устанавливаемым государством квотам (т.е. при изъятии, осуществляемом пропорционально их численностям и, следовательно, при формальном выполнении условия равенства охотничьей нагрузки на юге и севере), "ступень" неизменно будет возникать.
События, наблюдаемые на макропопуляционном уровне, "воплощаются в жизнь" на уровне микропопуляций. В свою очередь, микропопуляции не находятся в идентичных условиях: где-то пресс охоты выше, где-то меньше. И не только из-за того, что в каких-то местах браконьерство более распространено, а в каких-то менее. Просто охотники не распределены равномерно по пространству. В силу известных причин они тяготеют к тем участкам охотничьих угодий, до которых можно добраться без особого труда. Отсюда можно ожидать, что микропопуляции, занимающие территории вблизи транспортных магистралей и крупных населенных пунктов, в целом будут менее "весомы", чем микропопуляции из малодоступных и малонаселенных мест.
Это ожидание легко проверить - нанесением на областные карты средних масс для каждого из районов. После выполнения такой процедуры выяснилось, что лоси, обитающие ближе к "цивилизованным" местам легче на 4-6%. Таким образом, даже использование столь грубого (но единственно возможного) подхода, как порайонное деление, позволило обнаружить различия.
Итак, и на микро- и на макроуровне выявляется эффект избирательности изъятия. Об этом эффекте различные авторы писали много, но почти всегда имели ввиду избирательность по возрасту. То есть, масса лосей падает (как большинство из них считало и считает) только из-за того, что охотник, выбирая более крупное, а, следовательно, более взрослое животное, смещает возрастное распределение популяции в сторону ее омоложения, а значит и более легкой массы.
Заметим, что все те проценты, которые мы приводили выше, никак не связаны с избирательностью по возрасту.
Мы рассматривали только результат действия "чистого" искусственного (наводимого человеком) отбора, который оставляет след в следующем поколении в виде изменения генофонда и действует внутри отдельных возрастных классов. Тем не менее, интересно было указать, какая доля при избирательном изъятии в изменении массы (рассчитанной "скопом" для всех возрастных групп) приходится на возрастную структуру, а какая на действие "чистого" отбора.
Оказалось, что как на микро-, так и на макропопуляционном уровне, изменение возрастной структуры обеспечивает всего лишь 30-50% изменения "средневозрастной" массы, остальное приходится на последствия отбора внутри отдельных возрастных классов, т.е. на то, что закрепляется в популяциях и выявляется как 10 и 4-6 % различия между ними (различия с учетом возрастной структуры, естественно, гораздо выше этих процентов в полтора — два раза).
Следовательно, Европейский лось представлен по существу тремя группировками: южной, переходной и северной. Оптимальна для обитания лося переходная зона. Она характеризуется наибольшей плотностью и наименьшей плодовитостью, что характерно для успешных группировок. Все группировки отличаются морфологически, а также этологически (по проценту мигрантов) – Рожков и др., 2001.
В таблице 2 кратко охарактеризованы все три основные подразделения (группировки).
Недавно нами были начаты исследования (Холодова и др., 2005 - в этой статье см. подробно изложенную методику) по изменчивости нуклеотидных последовательностей гипервариабельного участка контрольного региона (левый домен) митохондриальной (мт) ДНК (т.е. выявлялось популяционное разнообразие гаплотипов
Таблица 2
Основные параметры, характеризующие три главных подразделения
Европейской макропопуляции
Параметры | Северная макропопуляция(тайга) |
Переходная зона |
Южная макропопуляция |
Масса туши | около 165 кг | переход от 165 до 140 кг | около 140 кг |
Эмбрионов на самку | 0,98 | 0,87 | 1,01 |
Плотность х1000 | 0,6-1,0 | 2,0-4,0 | около 0,2 - 0,5 |
% мигрантов | 70-80 | 0-5 | 50-60 |
На европейской территории РФ мы обнаружили 14 гаплотипов, из которых пять основных и 9 уникальных. В таблице 3 сгруппированы данные по их принадлежности к той или иной популяции.
Таблица 3
Распределение основных гаплотипов мтДНК лосей европейской части РФ
Популяция | n | Частота гаплотипа | ||||
L1 | L2 | L14 | L46 | L84 | ||
Южная макропопуляция (Рязанская, Тульская, Калужская, Московская, Владимирская) | 10 | 0,90 | 0,10 |
|
|
|
"Переходная» макропопуляция (Тверская, Ярославская, Костромская) | 29 | 0,48 | 0,45 | 0,07 |
|
|
Северная макропопуляция (Вологодская, Новгородская, Ленинградская Карелия, Архангельская) | 37 | 0,27 | 0,62 |
| 0,11 |
|
Ленинградская мезо-популяция (Ленинградская, Карелия) | 17 | 0,18 | 0,59 |
| 0,23 |
|
Популяционные группировки Архангельской области | 8 | 0,13 | 0,87 |
|
|
|
Скандинавская макропопуляция (Мурманская) | 3 | 0,33 |
|
|
| 0,67 |
В южной, "переходной" и северной макропопуляциях абсолютно доминируют два гаплотипа L1 и L2. Остальные (L14, L46) встречающиеся с небольшой частотой только в двух группировках, а так же большинство уникальных гаплотипов, близки по нуклеотидным последовательностям либо к L1, либо к L2, представляя собой по сути дела варианты этих гаплотипов. На юге частота гаплотипа L1 много больше частоты L2, в "переходной" зоне частоты L1 и L2 приблизительно равны и на севере частота L2 значительно выше L1. Если данный результат не следствие ошибки выборки (которая значительна) или следствие недостаточного количества охваченных исследованиями территорий, то мы явно имеем дело с изменяющимся с севера на юг градиентом частот, т.е. с подобием того, что мы только что рассмотрели на морфологическом уровне.
Территорию Скандинавии населяет лось несущий другой набор гаплотипов значительно отличающихся от L1 и L2 (Hundermark et al., 2002). "Скандинавские" гаплотипы выявлены нами в Мурманской области (L84). Единственный уникальный гаплотип обнаруженный на этой территории, так же принадлежит к скандинавской "гаплогруппе" являясь вариантом L84.
Ранее мы уже отмечали, что Мурманскую область и север Карелии, населяет отдельная, мезопопуляционная группировка, по-видимому, входящая в состав северной макропопуляции (Рожков и др., 2001). Генетические данные требуют небольшой корректировки. Они свидетельствуют о том, что эта мезопопуляция является частью скандинавской макропопуляции, которая, скорее всего, охватывает не только Швецию, но и Норвегию и север Финляндии (рис. 3)
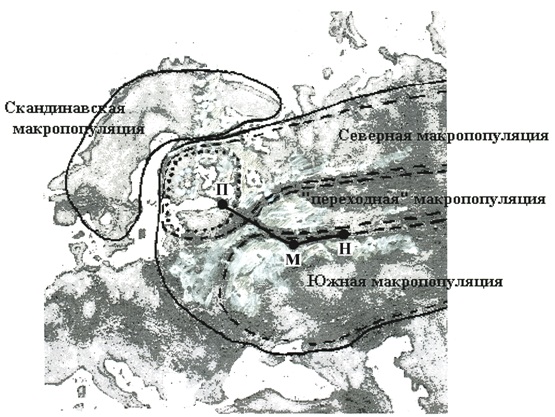
Рис. 3. Границы популяций лося на территории Европы.
Сплошные линии – границы макропопуляций высокого ранга (скандинавский и восточно-европейской).
Прерывистые линии – границы северной, южной и "переходной" макропопуляций, имеющих более низкий ранг.
Пунктир – границы ленинградской мезопопуляции.
П – Санкт-Перербург, М – Москва, Н – Нижний Новгород. Отрезок П – М – завершаемая ж/д трасса, М – Н – планируемая.
Южная, по-видимому, центральная часть Карелии, а также вся Ленинградская область занята другой мезопопуляцией входящей в состав северной макропопуляции о чём одинаково свидетельствуют как данные по фенотипу (Рожков и др., 2001), так и генетические данные. Её отличительной особенностью является наличие гаплотипа L46, близкого, как мы отмечали, по структуре к L1 и встречающегося как в Карелии, так и в Ленинградской области. Подобные гаплотипы характерны и для Финляндии, что позволяет продолжить границы Ленинградской мезопопуляции на южную и центральную части этой территории (на рисунке 7 мезопопуляция выделена пунктиром).
Следует отметить, что в целом результаты генетических исследований подтверждают то, что было ранее выявлено при анализе фенотипической изменчивости (Рожков и др., 2001). Однако, эти параллели между "генетикой" и "морфологией" ставят новые вопросы. Все особенности "морфологии" северных, южных и "переходных" лосей видимо сформировались в основном под действием отбора. Не свидетельствует ли это о том, что и наблюдаемая генетическая картина так же результат его действия.
Обычно считается, что генетическая изменчивость подобного типа нейтральна (или почти нейтральна) и, следовательно, её преобразования связаны лишь с объединённым действием генетического дрейфа (понимаемого в широком смысле слова) и мутационного процесса.
Тем не менее, следует, по-видимому, признать, что ни принцип основателя, ни предполагаемые катастрофические снижения численности, вряд ли способны дать в данном случае удовлетворительное объяснение. Южная макропопуляция сформировалась за срок меньший, чем 100 лет за счёт выселенцев из "переходной" зоны. В значительной степени, по-видимому, за счёт той же "переходной" зоны формировалось в отдалённом прошлом и население севера, так как эта зона вряд ли всегда была равно пригодной для обитания лося. Периодически повторяющиеся выселения из "переходной" зоны десятков особей, наверняка исключали заметное действие эффекта (принципа) основателя (к тому же этот эффект, как, впрочем, и другие формы генетического дрейфа, не приводит к закономерно меняющимся частотным градиентам). Видимо, проще предположить, что изменчивость гипервариабельного участка контрольного региона мтДНК далеко не нейтральна. В центральной, наиболее оптимальной для жизни лося "переходной" зоне, "уживаются" оба гаплотипа (L1 и L2), а в субоптимальных зонах преимущество получает на юге L1, а на севере L2. То есть, мы полагаем, что отбор шёл (а может быть идёт и поныне) на севере и на юге в противоположных направлениях, т.е. подобно тому, как он действует по морфологическим признакам.
Как мы отмечали выше, ареал переходной макропопуляции охватывает территорию с наиболее оптимальными для лося условиями и именно здесь наиболее высока его численность и только здесь он практически не повержен риску вымирания (конечно, если этому прямо не поспособствует человек). Эта зона по крайней мере на протяжении исторического времени (а скорее всего на протяжении 10 тыс. лет – окончание ледникового периода) вряд ли существенно меняла свое местоположение и являлась "питающей" для севера и юга.
То есть при исчезновении (вследствие вымирания) лося на севере (умеренно стабильная зона) и/или юге (не стабильная зона) обе эти территории вновь заселялись из переходной зоны. Происходило это за счет выселения из нее в основном, по-видимому, молодых (наиболее подвижных у многих видов) особей и образования на восстановившихся после экологических кризисов территориях новых популяционных группировок.
Таким образом, длительно, абсолютно самостоятельно (без "подпитки) южная и отчасти северная макропопуляционные группировки существовать не способны, так как находятся в экстремальных и субэкстремальных для обитания вида зонах. Правда на севере даже в периоды экологических кризисов возможно сохранение собственных очагов переживания вида, из которых также может начаться восстановление макропопуляции. Это обстоятельство позволяет рассматривать северную часть ареала вида как много более благополучную (сравнительно с южной) территорию.
Для юга сохранение очагов переживания более, чем проблематично, а скорее всего не возможно, так как и в относительно благоприятные годы площади лесных массивов (южнее центральной части Тульской и других прилегающих к ней областей) способны поддерживать только эфемерные популяционные группировки, которые не способны к длительному самостоятельному существованию.
В настоящее время заканчивается строительство скоростной трассы Москва – Санкт-Петербург (рис.3). Она проходит как раз по границе между южной и переходной макропопуляциями. На всем своем протяжении трасса изолирована защитными ограждениями, не преодолимыми для животных. Потенциально преодолимы лишь участки под мостами, и то только в зимнее время. Однако перемещения животных в первой половине зимы затухают, их миграционные пути никак не связаны с местонахождениями мостов на реках (хотя и можно предположить более вероятное движение особей вдоль рек, обильно поросших ивой), а сами по себе заграждения могут играть отпугивающую роль (тем более вблизи охраняемых железнодорожных мостов).
Отсюда возникает опасность изоляции южной макропопуляционной группировки от подпитывающей ее переходной зоны. Пока это касается только западной части ареала. В будущем планируется "отделить" переходную зону от южной макропопуляции и на востоке скоростной железнодорожной трассой Москва – Нижний Новгород (рис. 3). В этом случае будет значительно перекрыт переход лосей с севера на юг, в результате чего ближайший экологический кризис может привести к исчезновению южной макропопуляции, что ранее уже имело место (Данилкин, 2001) и к невозможности ее быстрого последующего восстановления.
__________________________________
1Столь широкие пределы определяются экологическими различиями территорий и тем, находится ли группировка в депрессии или "расцвете".
2Имеется ввиду не только законная охота, но и браконьерство.
3При трофейной охоте особи с наиболее массивными рогами, степень развития которых прямо скоррелирована с массой животных.
4Гаплотипы в некотором смысле подобны аллелям ядерных генов, но в отличие от последних гаплотипы "одновариантны" – каждый гаплотип несет только один вариант нуклеотидных последовательностей.
Литература
Давыдов А.В., Пискунов О.Д., Проняев А.В., Рожков Ю.И. Пространственная дифференциация лося Евразии (Alces alces L.) по результатам оценки охотничьих трофеев // Вестник охотоведения, 2004, т. 1, № 1, С.36-40.
Данилкин А.А. Оленьи //Млекопитающие России и сопредельных регионов. М.: ГЕОС, 1999. С. 301-358.
Рожков Ю.И., Проняев А.В., Пискунов О.Д., Овсюкова Н.Э., Давыдов А.В., Рожкова Л.В. Лось. Популяционно-биологический анализ лицензионной информации // Охотничьи животные России. Вып. 4. М.: Центрохотконтроль, 2001. 263 с.
Холодова М.В., Давыдов А.В., Мещерский И.Г., Пискунов О.Д., Рожков Ю.И. Изучение молекулярно-генетического разнообразия лося (Alces alces L.) центральный и северо-западной части России: анализ мт ДНК // Вестник охотоведения, 2005. т. 2, № 1. С. 26-33.
Hundermark K.J., Shields G.F., Udina I.G., Bowyer R.T., Danilkin A.A., Schwartz C.C. Mitochondrial phylogeography of Moose (Alces alces L.): late Pleistocene divergence and population expansion // Molec.Phyl. and Evol. 2002. V.22, № 3. P. 375-387.